אמבריולוגיה (Embryology, מיוונית :ἔμβρυον; 'זה שעוד לא נולד') היא המדע העוסק בהתפתחות העובר, מרגע הפריית הביצית ועד ללידה. תהליך ההפריה מציין את האיחוד שבין תא זרע לבין ביצית, ובסיומו מתקבלת זיגוטה. לאורך תקופת ההיריון משתנה הזיגוטה מאורגניזם חד-תאי לאורגניזם רב-תאי.
תהליך ההפריה (Fertilization) מתייחס לחיבור שבין תא זרע לבין ביצית. במהלך ההפריה ראשו של הזרע, האקרוזום, נפרץ ותכולתו נשפכת החוצה. תהליך זה מוביל לפגם במעטפת החלבונית של הביצית, ה-zona pellucida. דרך החריר שנוצר מטענו הגנטי של הזרעון חודר לביצית, ולבסוף מתאחד עם המטען הגנטי הנשי.
התפתחות העובר מחולקת באופן קטגורי לשתי תקופות:
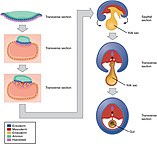
השלב האמבריוני (Embryonic stage) - שלב התפתחותי מוקדם, אשר בבני אדם כולל את 8 השבועות הראשונים מיד לאחר ההפריה. במהלך התקופה האמבריונית תהליך בידול התאים מואץ, מערכות תמיכה לתאים נוצרות ונוצרים לראשונה כלל איברי ומערכות הגוף. השלב מתחיל כאשר הבלסטוציט מתחבר לדופן הרחם ונקרא עובר. שלוש שכבות של תאים נוצרות. השכבה הפנימית של התאים נקראת אנדודרמה, אשר תתפתח למערכת העיכול ומערכת הנשימה. המזודרמה היא השכבה האמצעית, אשר תתפתח למערכת הדם, עצמות, שרירים, מערכת ההפרשה ומערכת הרבייה. השכבה החיצונית נקראת אקטודרמה או אפידרמיס שתתפתח למערכת העצבים והמוח, קולטנים חושיים (אוזניים, אף, ועיניים לדוגמה) וחלקי עור (כגון שיער וציפורניים). כל חלק בגוף מתפתח משלוש השכבות האלה. כאשר שלוש השכבות נוצרות אצל העובר, מערכות תומכות חיים (שק השפיר, חבל הטבור והשליה) מתפתחות במהירות. .
השלב הפיטאלי (Fetal stage) - שלב התפתחותי מאוחר, מן השבוע ה-9 ועד ללידה. בשלב זה איברי הגוף השונים כבר נוצרו, ומעתה ואילך מתחילה הבשלתם. התפתחות האיברים איננה מסתיימת בלידה, ולמעשה במשך כל תקופת החיים איברי הגוף משתנים. דוגמה לכך היא התפתחות הריאות, אשר אורכת בממוצע כ-8 שנים בבני אדם.
אף על פי שהמונח "עובר" מתייחס להתפתחות הזיגוטה לאורך כל תקופת ההריון, ישנו פער התפתחותי גדול בין האמבריו (Embryo) לבין הפיטוס (Fetus).
הזרעון (בלעז; Spermatozoon) הוא תא הפלואידי, מאורך, ובעל כושר תנועה. לזרעון שלושה אזורים אנטומיים מרכזיים:
ראש - מבנה עגול אשר מכיל את האקרוזום, גרעין התא ומרבית האברונים התאיים.
צוואר - מבנה צר וארוך, אשר מכיל את כלל המיטוכונדריה הזכריות.
זנב - מבנה מאורך אשר אחראי על יכולת התנועה של התא. מבחינת ביולוגיה של התא מבנה זה נקרא שוטון (פלג'לום)[1][2].
האקרוזום הוא שקיק ממולא באנזימים פרוטאוליטיים, אנזימים בעלי כושר חיתוך חלבונים. במהלך ההפריה, הזרעון מפריש את תוכן האקרוזום, ובכך פוגע במעטפת החלבונית של הביצית - Zona pellucida.
התפתחותו של תא הזרע מתחילה באשכים. תנועתיות הזרעון היא תכונה נרכשת, אשר מופיעה לראשונה רק מחוץ לאשך, כאשר הזרעון מועבר לאפידידימיס. התהליך בו הזרעון מקבל את כושר תנועתו נקרא מטורציה (Maturation). לאחר השפיכה, תאים אלו עוברים תהליך התבגרות נוסף שנקרא קפסיטציה (Capacitation), אשר בסופו מתקבלת תכונה נוספת - הפריה. במהלך הקפסיטציה הזרעונים הופכים להיפרמוטיליים (בעלי כושר תנועה מוגבר) ויציבות מעטפת האקרוזום פוחתת. מאפיינים אלו מסייעים לזרעון לנוע אל עבר הביצית, ולהפריש את תוכן האקרוזום על ה-Zona Pellucida.
יצירת תאי הזרע מתחילה במהלך גיל הבגרות הזכרית. מסלול החיים של הזרעון הוא כשלושה שבועות.
תאי סרטולי הם מולטי-מורפולוגיים אשר תומכים תהליך ההתפתחות של תאי הזרע - ספרמטוגנזה. כמו כן, תאים אלו מחוברים זה לזה ב-tight junctions וב-gap junctions, כך שהם יוצרים הפרדה מוחלטת ורציפה בין המתחם בו תאי הזרע מתפתחים, לבין שאר תאי הגוף. היצמדות זו מכונה בשם מחסום דם-אשך (Blood-Testis Barrier). באופן עקרוני, מחסום זה דומה למחסום דם-מוח (Blood-Brain Barrier).
תאי ליידיג הם תאים גדולים הממוקמים מחוץ למחסום דם-אשך. תאים אלו אחראיים על ייצור והפרשת הורמון המין הזכרי טסטוסטרון.
הביצית היא תא סימטרי, עגול וגדול למדי. הביצית נוצרת בשחלה. בניגוד לתאי הזרע, הביצית נוצרת כבר במהלך ההתפתחות העוברית. יתרה מכך, היווצרות הביציות פוסקת גם היא במהלך התקופה העוברית, ולא נוצרות ביציות חדשות במהלך החיים. בעת הלידה, לאישה ממוצעת ישנן כ-2 מיליון ביציות. אף על פי כן, רק כ-40,000 ביציות ישרדו עד גיל הבגרות, ובמהלך החיים רק כ-400 ישלימו ביוץ.
ניגוד בולט בין תא זרע לבין תא ביצית הוא הפלואידיות. בעוד שתא זרע מתפתח באופן רציף ועוזב את האשך כתא הפלואידי מושלם, אין זה כך עבור הביציות. במשך כל תקופת החיים הביציות תקועות במהלך חלוקה מיוטית. בלידה, כלל הביציות הן בשלב פרופאזה 1. במהלך גיל הבגרות, כאשר המחזור הנשי מתחיל, בכל חודש ביצית אחת משלימה את החלוקה המיוטית הראשונה, פורצת מתוך השחלה לחלל הבטן, ומתחילה את החלוקה המיוטית השנייה. תהליך זה נקרא ביוץ (Ovulation). לאחר מכן הביצית נקלטת על ידי החצוצרות, וחודרת לתוכן. החלוקה המיוטית איננה מגיעה לסיומה במהלך הביוץ, אלא נעצרת במטפאזה 2.
כאשר ישנו תא זרע שעבר קפסיטציה בסביבה, עשויה להתרחש הפריה. רק לאחר ההפריה מסתיימת החלוקה המיוטית השנייה, ומתקבלת ביצית הפלואידית. החלוקה המיוטית מושלמת תוך כדי חדירת הפרו-נוקלאוס הזכרי, ולכן הביצית איננה נחה באינטרפאזה לעולם. כל עוד אין זרעון זמין בסביבה, הביצית תמות והמחזור החודשי יתחדש.
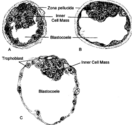
זקיק שחלתי בוגר, בעל שכבות רבות של תאי גרנולוזה. ניתן לראות את הביצית, מוקפת ב-Zona Pellucida (בתמונה נקרא Zona Striata), ולאחריה תאי קומולוס. גרעין הביצית נקרא Germinal Vesicle, לציון העובדה שהביצית נחה בפרופאזה 1 (חלוקה מיוטית ראשונה).
בדומה לזרעונים, אשר נתמכים על ידי שני סוגי תאים; סרטולי וליידיג, גם הביצית נתמכת על ידי שני סוגי תאים מרכזיים; תאי גרנולוזה ותאי טקה (Theca cells). זקיק שחלתי (Ovarian Follicle) הוא מבנה אנטומי מורכב הכולל גם את הביצית, וגם את תאי התמך האישיים שלה.
תאי גרנולוזה מקיפים את הביצית במהלך התפתחותה. תאים אלו קשורים זה לזה ב-Tight junctions וב-Gap junctions, והם מוקפים במעטפת חלבונית קשיחה שנקראת 'השכבה הבזאלית' (Basal Lamina). צמתים אלו מבודדים באופן הרמטי את סביבת הביצית משאר תאי הגוף. בידוד תאי זה נקרא מחסום דם-זקיק (Blood-Follicle Barrier). השכבה הבזאלית מפרידה בין הזקיק לבין תאי הסטרומה השחלתיים.
ככל שהזקיק גדל ומתבגר, מספר שכבות תאי הגרנולוזה גדל גם הוא. התאים אשר ממוקמים בשכבה הפנימית ביותר, זו הצמודה לביצית עצמה, נקראים תאי קומולוס (Cumulus cells). בין תאי הקומולוס לבין הביצית מצטברת מעטפת חלבונית אשר מגנה על הביצית. מעטפת זו נקראת Zona Pellucida.
תאי טקה מתפתחים מתאי סטרומה אשר מקיפים זקיקים בוגרים. בהמשך תאי הטקה מתרבים, ומתפצלים לשתי שכבות נפרדות - פנימית וחיצונית. השכבה הפנימית (Theca Interna) מקבילה בתפקודה לתאי הליידיג. תאים אלו מפרישים אסטרוגן, הורמון מין נשי.
התפתחות הזקיקים אורכת זמן רב (כמספר שבועות, עד כדי חודשים) והיא תלויה בעיקרה בוויסות אנדוקריני. ההורמונים המרכזיים אשר משפיעים על התבגרות הזקיקים הם FSH ו-LH. בכל חודש, הזקיק הגדול ביותר נבחר לעבור ביוץ, והביצית אותה הוא מכיל תחרוג מן השחלה אל עבר החצוצרות.
הפריה היא מאורע מרכזי בתחום האמבריולוגיה. תהליך זה אחראי באופן ישיר ליצירה של יצור חי חדש. במהלך ההפריה מתרחשים שינויים מרחיקי לכת בביצית, כאשר ניתן למנות ביניהם:
השלמת החלוקה המיוטית השנייה
הפרשת חומרים אשר משמידים זרעונים
מעבר מתא הפלואידי ממויין לתא דיפלואידי טוטיפוטנטי
אובדן יכולת ההפריה
הרס המיטוכונדריה הזכריות
לאחר הביוץ, הביצית נחה בחצוצרות במטפאזה 2, עטופה ב-zona pellucida. זרעון אשר השלים קפסיטציה נצמד ל-zona pellucida, חודר דרך ממברנת הביצית, וגורם להפריה.
היצמדות הזרעון ל-zona pellucida מובילה לשחרור האנזימים הפרוטאוליטיים אשר מאוחסנים באקרוזום. שחרור זה נקרא תגובה אקרוזומלית (Acrosome Reaction). אנזימים אלו מעכלים את חלבוני ה-zona pellucida (בבני אדם אתר המטרה הוא ככל הנראה ZP-2), ומערערים את יציבותה. לאחר חדירת מעטפת חלבונית זו, הזרעון נצמד לממברנת הביצית. זיהוי הקולטנים האחראיים להפריית הביצית הוא נושא אשר עומד בחזית המחקר מזה שנים רבות. אף על פי שנותר ערפל רב בתחום זה, מסתמן כיום כי צמד הליגנד-קולטן המרכזי אשר מקיים הפריה הואJuno-Izumo, כאשר Izumo ממוקם על גבי הזרעון, ואילו הקולטן Juno על גבי הביצית[3]. זיהוי המולקולות אשר אחראיות להפריה עשוי לשנות בצורה דרמטית את תחום הפוריות מחד, ואת מניעת ההריון מאידך.
צימוד הזרעון לביצית גורם להשלמת החלוקה המיוטית. פעולה זו מתאפשרת באמצעות עלייה חדה בריכוז הסידן, אשר מתפקד כמוסרן שניוני. ריכוז הסידן משתנה בעקבות החדרת אנזים זכרי - PLCζ.
כמו כן, האוסילציות בריכוז הסידן מובילות לשינויים מהותיים ב-zona pellucida. שקיקים מוכנים מבעוד מועד מופרשים מעבר לממברנת התא, ותוכנם מתאחד עם ה-zona pellucida, מחזק ומעבה אותה. שקיקים אלו נקראים גם גרנולות קורטיקליות, ולכן לתהליך הפרשה השקיקים נקבע הכינוי תגובה קורטיקלית (Cortical Reaction). מטרת התגובה הקורטיקלית היא מניעת פוליספרמיה (הפריות מרובות).
כלל התאים באורגניזם מולטי-סלולרי (רב-תאי) מכילים בגרעינםמטען גנטי זהה (מלבד במקרה של מוטציות או מקרים מיוחדים נוספים). עם זאת, ישנו פער מהותי בין תא עצם, לבין תא שריר או תא עור. פער זה נובע מכך שהתאים הללו מבטאים גנים שונים. התהליך בו תא מסוים משתנה, ומבטא גנים שונים, נקרא התמיינות תאית (Cellular Differentiation). מטרה בולטת הן באמבריולוגיה, והן בביולוגיה התפתחותית, היא חקר הגורמים להתמיינות תאית באורגניזמים שונים.
המחשה כיצד מפל ריכוזים עשוי להשפיע על אוסף של תאים. כתלות בכמות החומר המופרש (מסומן בסגול), התאים המקוריים יתמיינו לתאי מטרה שונים. ייתכן שחלק מן התאים לא ייחשפו כלל, וישנם תאים אשר יושרו בריכוז גבוה.
מקורם של כל תאי הגוף, בכל יצור חי, הוא הזיגוטה. תא זה הוא תא טוטיפוטנטי, כלומר, תא בעל יכולת התמיינות אין-סופית. לאחר היווצרותה, הזיגוטה מתחילה בחלוקה מיטוטית נרחבת. ריבוי התאים טומן בחובו את פוטנציאל ההתמיינות. כאשר העובר העתידי ייחשף לחומר מסוים, ייווצר גרדיאנט ריכוזים לאורך העובר עצמו. התאים הקרובים למקור יושפעו אחרת מתאים אשר ייחשפו לריכוז נמוך, הרחק ממקור ההפרשה.
ישנו מספר מצומצם של ווסתי התמיינות מולקולריים. כל ווסת מולקולרי נקשר לקולטן מתאים, ומהווה טריגר למסלול מעבר אותות, תהליך שנקרא Signal transduction. לכל מסלול תוצאה ייחודית לו, שיכולה להתבטא הן ברמה הגנטית, הן ברמה הטרנסקריפטומית, והן ברמת החלבונים.
ריכוז החומרים, תזמון הפרשתם, וסוג החומרים קובע בסופו של דבר לאיזה תא ספציפי יתמיין תא האב. באמצעות עקרונות אלה ניתן כיום למיין באופן מלאכותי תאים בתרבית.
להלן החומרים המרכזיים אשר אחראיים לדיפרנציאציה תאית:
זיגוטה צעירה. ניתן לראות את התקרבות הפרו-נוקלאוס הזכרי והפרו-נוקלאוס הנקבי. במילים אחרות, אלו הם הרגעים האחרונים של ההפריה, ובקרב יאוחד המטען הגנטי הזכרי והנקבי.
בבני אדם, השלב האמבריוני כולל את 8 השבועות הראשונים. תקופה זו מתאפיינת בשינויים רבים ובהתפתחות מהירה.
זיגוטה - תא דיפלואידי אשר נוצר מהפריית הביצית. הזיגוטה מתחילה בחלוקה מיטוטית מספר שעות לאחר היווצרותה. התאים הנוצרים בעקבות זאת נקראים בלסטומרים (Blastomeres).
מורולה עטופה בשכבה חלבונית הנקראת zona pelluca.מורולה (תותית) - צבר של 12–32 בלסטומרים[4].
בלסטוציסט. ניתן לראות את החלוקה לשכבה חיצונית - טרופובלסט, ולשכבה פנימית - אמבריובלסט (בתמונה האמבריובלסט מכונה בראשי התיבות ICM - Inner cell mass).בלסטולה - שבירת המעטפת החלבונית של הביצית (zona pellucida) מתרחשת כ-4 ימים לאחר ההפריה. הפריצה דרך מעטפת זו מאפשרת למורולה להתרחב. כמו כן, נוזלים ממלאים את המורולה ומפרידים אותה לשתי שכבות שונות; שכבה היקפית בשם "טרופובלסט"(Trophoblast), ושכבה פנימית, אשר תיצור את העובר העתידי, בשם "אמבריובלסט" (embryoblast). בשלב זה נקרא העובר בלסטוציסט (Blastocyst). במהלך היום ה-5 הבלסטוציסט עובר השרשה (Implantation) בדופן הרחם. לאחר ההשרשה, השכבה הפנימית של הבלסטוציסט מתמיינת לשתי שכבות נוספות; אפיבלסט (Epiblast)והיפובלסט (Hypoblast). שכבות אלו נעות במרחב ויוצרות צורה דיסקית.
גסטרולה מושרשת. ניתן לראות בבירור את החלוקה לשלוש שכבות הנבט; בצהוב - אנדודרם, באדום - מזודרם, ובכחול - אקטודרם.גסטרולה - במהלך השבוע השלישי לחיים דיסקה זו מתמיינת מחדש ל-3 שכבות נבט; אנדודרם, מזודרם ואקטודרם. תהליך זה חשוב מאין כמוהו בהתפתחות העוברית. התהליך בו הדיסקה הבילמינרית (דו-שכבתית) משתנה לשכבה טרילמינרית (תלת-שכבתית) נקרא גסטרולציה.
קיפול עוברי לאחר הגסטרולציה, סוף השבוע השלישי לחיים.נוירולה - בהמשך ההתפתחות, הדיסקה התלת-שכבתית משנה את צורתה לכדי צינור. שינוי צורני זה נקרא קיפול עוברי (Embryonic folding). במקביל לקיפול העוברי, שכבת הנבט החיצונית של הגסטרולה (האקטודרם) גם היא מתקפלת ויוצרת צינור בתהליך שנקרא נוירולציה (Neurolation). תהליך זה יוצר הלכה למעשה את מערכת העצבים בבעלי חוליות. בהמשך מתפתחים כל שאר איברי הגוף.
מן השבוע ה-9 ואילך, העובר מקבל צורה דומה לאדם בוגר. בשלב זה העובר נקרא פיטוס (Fetus). ב-31 השבועות שלאחר מכן איברי הגוף גדלים ומתחזקים. תהליך התפתחות האיברים נקרא הבשלה.
במהלך החודש הראשון לאחר הלידה, הוולד נקרא יילוד. במהלך תקופה זו ישנם שינויים מרחיקי לכת מבחינת מערכת הלב ומערכת הנשימה. כאשר ההתאמות הפיזיולוגיות מושלמות כהלכה, סיכויי השרידות להמשך ההתפתחות נחשבים לטובים. עם זאת, התקופה הנאונטאלית נחשבת לרגישה מבחינת הסכנה לחיים, בייחוד מפני מחלות נשימתיות ומחלות זיהומיות.
לאחר גיל חודש ועד לגיל שנה, היילוד נקרא תינוק. תקופה זו מתבטאת בגדילה מהירה. לרוב עד גיל שנה, משקל התינוק מוכפל פי 3 ממשקל הלידה, אורכו מתארך בכ-50% ומופיעות 6–8 שיני חלב.
תקופה המתבטאת בהשלמת כושר הפיריון. ביטויים להשלמת הבשלת הפוריות כוללים שיער ערווה, ניצני חזה, הרחבת האגן, שינוי קול ועוד. תקופת ההתבגרות מסתיימת עם התגרמות העצמות ועצירת התארכותן. ההתפתחות המוחית ממשיכה גם לתוך תקופת הבגרות.
מחזור האישה במספר רמות; רמת השחלה, טמפרטורה בזאלית, פעילות הורמונלית ושינויים רחמיים. מחזור האישה נספר מרגע הדימום. כשבועיים לאחר הדימום מתרחש הביוץ (ovulation), ובצמוד לאירוע זה מתרחשת ההפריה.
ישנו פער בין שבוע היריון (נקרא גם 'שבוע קליני') לבין שבוע ההתפתחות העוברית. פער זה נובע מכיוון שלא ניתן לדעת בוודאות מהו רגע ההפריה והיווצרות הזיגוטה. לעומת זאת, כן ניתן לתארך את הדימום הוסתי התקין האחרון. מסיבה זו נהוג למדוד את משך ההריון בשבועות קליניים ביחס לדימום הווסתי האחרון.
במחזור סדיר סטנדרטי, האורך 28 ימים, הביוץ מתרחש ביום ה-14, כשבועיים לאחר הוסת. לכן כאשר תיווצר הזיגוטה, אף על פי שזהו השבוע הראשון לחיים, האישה תחשב כבר בשבוע השלישי להריון.
מחזור נשי תקין איננו בהכרח אורך 28 ימים. למען האמת, מחזור תקין נע בין 21 ל-35 ימים. הביוץ מתרחש לרוב 14 יום לפני הדימום, ולא לאחריו. במילים אחרות, אישה בעלת מחזור סדיר האורך 35 ימים, ברגע ההפריה תחשב בשבוע קליני 4. אישה בעלת מחזור סדיר האורך 21 ימים, ברגע ההפריה תהיה בשבוע הקליני השני.
העיסוק בהבנת התפתחות הילוד האנושי ברחם אמו קסם לרבים מאנשי המדע, הפילוסופיה והרוח בתקופות שונות בהיסטוריה האנושית, החל מהתקופה שבה פרחה הפילוסופיה היוונית.
רבים פיתחו תאוריות, סברות ואגדות שונות אודות המתרחש ברחם האם ושלבי התפתחות היילוד.
האסכולה הפיתאגורית, אותה הנהיגו פיתגורס ופילולאוס, גרסה כי זרע העובר עבר מעין דגירה ברחם אמו במהלך שלבי התפתחותו.
על סמך אסכולה זו, לעובר שלושה מקורות:
חלקו הצמחי - מקורו בטבור; המתייחס לנפש הצומחת (הזנה).
האסכולה הרפואית של קוס (עירו של הרופא הנודע היפוקרטס), גרסה כי התפתחות העובר זהה לתהליך אפיית הלחם בתנור. לפי אסכולה זו כלל האיברים מתפתחים בעת ובעונה אחת.
בנוסף, סבר היפוקרטס כי ישנם שלבים מסוימים בהתפתחות העובר:
בסיום תקופת ארבעת החודשים הראשונים להיריון נוצר בשר העובר.
עיקר ההתפתחות העיונית והשיטתית לגבי האמבריולוגיה, בתקופה זו, נזקפת לזכותו של אריסטו, שתיאר את צינורות הטבור והגה את הרעיון כי איברי העובר עוברים תהליך התהוות ולאחר מכן תהליך התפתחות, וזאת בניגוד מוחלט לדעה הגורסת כי העובר מתחיל להתפתח כשמצויים בו כבר כל איבריו.
משרטוטיו ורישומיו של לאונרדו דה וינצ'י, המפתיעים בדיוקם. 1513 בקירוב.
על פי אבן סינא, פילוסוף ורופא ממוצא פרסי, שהושפע מגלנוס, העובר מתהווה משלוש שלפוחיות שונות שמהן מתפתחים הכבד, הלב והמוח.
על פי האנטומיה ההודית, כל האיברים מתפתחים בו זמנית משבעה קרומים שונים, כאשר לכל חודש בהיריון שלבים מיוחדים משלו בהתרקמות האיברים.
בימי הביניים הידיעות בדבר האמבריולוגיה היו מעורפלות מאוד, אך לאונרדו דה וינצ'י צייר לראשונה את העובר ברחם אמו, בדיוק רב ומרשים למדי ביחס לידיעות במאה ה-21.
במאה ה-17 חלה התקדמות רבה במדע האמבריולוגיה.
האנטומיסט והמדעןהרונימוס פבריציוס החל בהשוואת האמבריולוגיה של האדם להתפתחות בעלי חיים אחרים.
החוקרויליאם הארווי - מגלה מחזור הדם, העלה הנחה יסודית לפיה התפתחותם של יונקים ברחם אמם איננה שונה מהתפתחות בעלי חיים אחרים בביציהם.
במחצית השנייה של המאה ה-17, עם המצאת המיקרוסקופ ושילובו במחקר הרפואי על ידי מרצ'לו מלפיגי, חלה קפיצת דרך משמעותית במחקר האמבריולוגי. נתגלו שלפוחיות בשחלה וכן נעשו מחקרים בהתפתחות הביצית בשחלה ובחצוצרת הרחם של הפרה ובשלבים התפתחותיים מתקדמים יותר.
בנוסף, נתגלו תאים בשכבת הזרע האנושי בידי אנטוני ואן לוונהוק ותלמידו.
במאה ה-18 פרצה מחלוקת בין המחנה שצידד בגישה האבולוציונית (פרפורמציונית) שעימה נמנו ספלנצני והאלר ובין המחנה שצידד בתורת האפיגנזה שייסד ר. פ. וולף, שהושפע ממרצ'לו מלפיגי. המחלוקת השפיעה רבות על התפתחות האמבריולוגיה ונתנה דחיפה למדע זה.
המאה ה-19 היוותה המשך להתפתחותה של האמבריולוגיה. בתקופה זו הונח היסוד לאמבריולוגיה המשווה בידי אטיין ז'ופרואה סנט-אילר שהבחין בקווים משותפים בהתפתחות קבוצות בעלי החיים השונות.
במחצית השנייה של המאה ה-19 נפתח פתח להבנת התפתחות החולייתנים עם החידושים הטכנולוגיים בתחום ההיסטולוגי.
זו הייתה גם התקופה שארנסט הקל פיתח בה את החוק הביוגנטי האומר שההתפתחות האורגנית של יצור חי מהווה מעין תקציר של תולדות המין הטקסונומי שלו. אף על פי שהחוק חל באופן מסוים במקרים רבים, הוא אינו מקובל על הביולוגים בני ימינו. זו הייתה גם התקופה שקרל ארנסט פון באר (אנ') פיתח את חוקיו שטוענים בכל אחד משלבי ההתפתחות של עובר, הוא יהיה דומה לעוברים של בעלי חיים דומים ולא לפרט בוגר של אותו בעל חיים.
תחומי המחקר באמבריולוגיה רבים ומגוונים. בדומה למקצועות ביורפואיים נוספים, חזית המחקר כוללת הן מחקרי יסוד והן מחקרים קליניים. בין היתר ניתן למנות:
אמבריולוגיה תפקודית: תחום זה חוקר את כלל הגורמים המשפיעים על התפתחות תאי העובר, בדגש על כוחות פיזיקליים, סיגנלים ביוכימיים ופקטורי שעתוק. תחום זה מכונה גם 'פיזיולוגיה של התפתחותית', או 'מכניקה התפתחותית'. ראו ערך ביולוגיה התפתחותית למידע נוסף.
אמבריולוגיה מורפולוגית: מדע תיאורי העוסק במכלול השינויים העיקריים החלים בגופו של העובר, עד הגעתו להתפתחות מלאה. תחום זה מכונה גם 'אנטומיה התפתחותית', ולעיתים 'אמבריולוגיה ניסויית'.
טרטולוגיה (Teratology): מדע העוסק בגורמים לפגמים התפתחותיים והיווצרות מומים. האטיולוגיה (הגורם) למרבית המומים היא שילוב של אלמנטים גנטיים וסביבתיים. גורמי המוות המרכזיים בתקופת הילדות מקורם במומים.
Keith L. Moore & T.V.N Persaud & Mark G. Torchia. (2020). The Developing Human. Clinically Oriented Embryology. 11th ed. Elvesier. ISBN 978-0-323-61154-1.
Alberts. (2015). Molecular Biology of the Cell. 6th ed. Pages 940-942. Elvesier. ISBN 978-0-8153-4432-2.
עזרה זהר ויאיר שפירא, הגוף ותפקודו - אנטומיה, פיזיולוגיה, תזונה, הוצאת עם עובד, 2005, עמודים 118–137.
עדי מרקוזה-הס, דידה פרנקל ונורית בשן, ביולוגיה של האדם, הוצאת האוניברסיטה העברית 2002, עמודים 181–202.
^השימוש במונח "שוטון" מטעה. ישנו הבדל מהותי בין הפלג'לום האאוקריוטי לבין אחיו הפרוקריוטי, הן ברמה המולקולרית, הן ברמה המבנית והן ברמה התפקודית. השוטון האאוקריוטי בנוי מן החלבונים דינאין (Dynein) וטובולין (Tubulin). תנועתו היא כשל גל. לעומת זאת, החלבון אשר מרכיב את השוטון הפרוקריוטי הוא פלג'לין (Flagellin), ותנועתו היא כשל פרופלור. הסיבה לשימוש במונח שוטון מקורה בטעות היסטורית מצערת.
^Alberts & Johnson & Lewis & Morgan & Raff & Roberts and Walter, Molecular Biology of the Cell, Third Avenue, 711, New York, NY 10017, US: Garland Science, Taylor & Francis Group, LLC, 2015
^Bianchi E, Doe B, Goulding D, Wright GJ, Juno is the egg Izumo receptor and is essential for mammalian fertilization, Nature
^Keith L. Moore, The Developing Human - Clinically Oriented Embryology, Elvesier Inc@, 2020



 ערכים מורחבים – תא זרע, ספרמטוגנזה
ערכים מורחבים – תא זרע, ספרמטוגנזה





.svg)